


주메뉴
- About IBS 연구원소개
-
Research Centers
연구단소개
- Research Outcomes
- Mathematics
- Physics
- Center for Underground Physics
- Center for Theoretical Physics of the Universe (Particle Theory and Cosmology Group)
- Center for Theoretical Physics of the Universe (Cosmology, Gravity and Astroparticle Physics Group)
- Dark Matter Axion Group
- Center for Artificial Low Dimensional Electronic Systems
- Center for Theoretical Physics of Complex Systems
- Center for Quantum Nanoscience
- Center for Exotic Nuclear Studies
- Center for Van der Waals Quantum Solids
- Center for Relativistic Laser Science
- Chemistry
- Life Sciences
- Earth Science
- Interdisciplinary
- Center for Neuroscience Imaging Research (Neuro Technology Group)
- Center for Neuroscience Imaging Research (Cognitive and Computational Neuroscience Group)
- Center for Algorithmic and Robotized Synthesis
- Center for Genome Engineering
- Center for Nanomedicine
- Center for Biomolecular and Cellular Structure
- Center for 2D Quantum Heterostructures
- Center for Quantum Conversion Research
- Institutes
- Korea Virus Research Institute
- News Center 뉴스 센터
- Career 인재초빙
- Living in Korea IBS School-UST
- IBS School 윤리경영

주메뉴
- About IBS
-
Research Centers
- Research Outcomes
- Mathematics
- Physics
- Center for Underground Physics
- Center for Theoretical Physics of the Universe (Particle Theory and Cosmology Group)
- Center for Theoretical Physics of the Universe (Cosmology, Gravity and Astroparticle Physics Group)
- Dark Matter Axion Group
- Center for Artificial Low Dimensional Electronic Systems
- Center for Theoretical Physics of Complex Systems
- Center for Quantum Nanoscience
- Center for Exotic Nuclear Studies
- Center for Van der Waals Quantum Solids
- Center for Relativistic Laser Science
- Chemistry
- Life Sciences
- Earth Science
- Interdisciplinary
- Center for Neuroscience Imaging Research (Neuro Technology Group)
- Center for Neuroscience Imaging Research (Cognitive and Computational Neuroscience Group)
- Center for Algorithmic and Robotized Synthesis
- Center for Genome Engineering
- Center for Nanomedicine
- Center for Biomolecular and Cellular Structure
- Center for 2D Quantum Heterostructures
- Center for Quantum Conversion Research
- Institutes
- Korea Virus Research Institute
- News Center
- Career
- Living in Korea
- IBS School





News Center

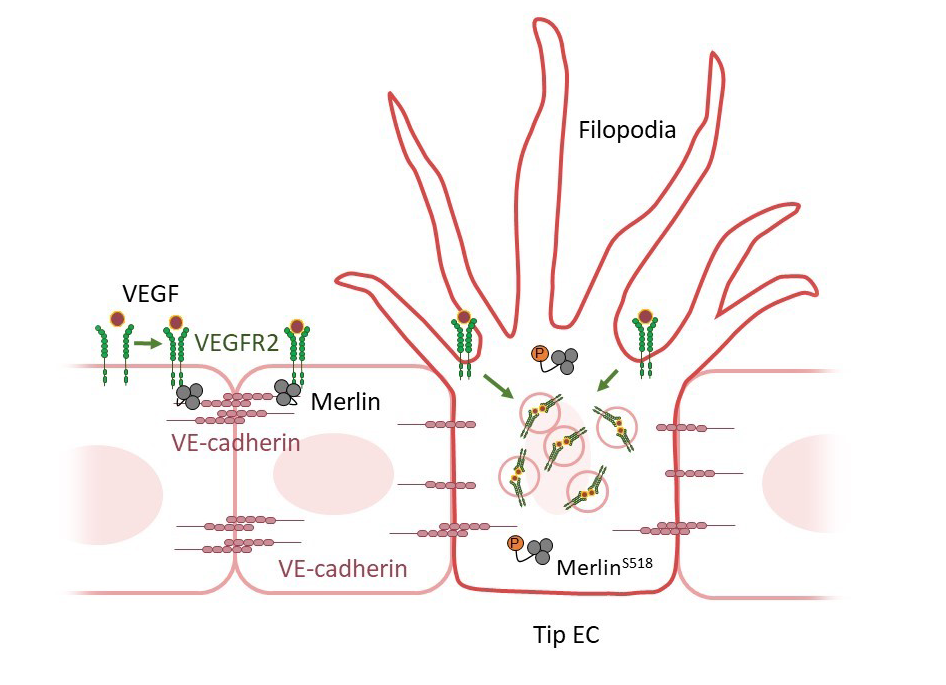
“Merlin” serves as a gatekeeper in new blood vessel formation- Merlin suppresses internalization of VEGFR2, which is important for the formation of ‘tip’ endothelial cells - Blood vessels are the body’s essential architecture that exists in order to nurture cells with sufficient nutrients and oxygen. Angiogenesis, or the formation of new blood vessels, is of great research interest in cancer medicine. As tumors need to generate new blood vessels to grow, inhibiting angiogenesis has been suggested as a promising approach to treating cancer. Sprouting angiogenesis is the process where new sprouts are formed from pre-existing vessels. The key biological process in sprouting angiogenesis involves balancing the formation of ‘tip’ and ‘stalk’ endothelial cells (ECs). This is a process called tip-stalk specification. Tip ECs are highly motile with many long and dynamic filopodia, whereas stalk ECs are highly proliferative with fewer filopodia. These functionally distinct tip and stalk cells coordinate into a branched network of vessels for sprouting angiogenesis. Morphologically, tip EC induction is the foremost step for sprouting angiogenesis. The formation of the tip ECs is induced by a pathway called vascular endothelial growth factor (VEGF)-A (given as VEGF hereafter) signaling. Molecularly, intracellular activation of VEGFR2 signaling is initiated by ligand-induced, clathrin-dependent VEGFR2 internalization. Scientists have long been curious how one particular EC ends up possessing higher VEGFR2 signaling activity than its neighbors and is thus fated to become a tip EC during sprouting angiogenesis. Led by Dr. BAE Jung Hyun and Professor KOH Gou Young, researchers at the Center for Vascular Research within the Institute for Basic Science (IBS) in Daejeon, South Korea discovered that a protein called Merlin is indispensable for tip EC induction by regulating intracellular VEGFR2 downstream signaling during sprouting angiogenesis. Merlin, encoded by the neurofibromatosis type-2 gene NF2, has been known for quite a while to function as a tumor suppressor. It is ubiquitously expressed in all types of cells and acts as a membrane-associated protein, which plays a gatekeeping role in signal transduction from membrane receptors in a cell density-dependent manner. Nevertheless, it has not been investigated whether endothelial Merlin is essential in sprouting angiogenesis and, in detail, which intracellular signaling is regulated by endothelial Merlin. The researchers showed that the density of the EC in the environment was the key factor. When the localization and density of Merlin in the cells under low or high culture density were compared, Merlin in the high cell density culture was able to better form complex with VEGFR2. The researchers showed that this is due to the fact that the presence of VE-cadherin, a cell-to-cell junction protein, is required for Merlin and VEGFR2 to form a complex with one another. This formation of the Merlin-VEGFR2 complex suppresses VEGFR2 internalization, thereby suppressing the pathway. In contrast, Merlin in a low-density ECs environment with relatively low VE-cadherin density allows a high level of VEGFR2 signaling and ERK activation, which leads to tip EC induction (Fig.1). The researchers investigated the importance of cell density on Merlin-VEGFR2 complex formation by studying Merlin in its normal form, inactive phosphorylated form (pMerlin (S518), short for phosphorylated Merlin at Serine 518), and a variant that cannot be phosphorylated. It was shown that Merlin engages in physical interaction with VEGFR2 and VE-cadherin in dense culture, which has a low pMerlin (S518), but Merlin does not engage in this interaction in sparse culture, with a high pMerlin (S518) in cultured ECs. Thus, the interaction between Merlin and VEGFR2 depends on pMerlin (S518), which is regulated by cell density. “The role of Merlin in EC fulfills the missing piece for sprouting angiogenesis. Sprouting angiogenesis should be regulated by the coordinated balance of promotion and inhibition of signaling through Merlin,” explains the first author, BAE Jung Hyun. The researchers further demonstrated that Merlin also plays a gatekeeping role in maintaining capillary integrity and proper angiogenesis by suppressing the unnecessary formation of filopodia and sprouting from the adult vessels in normal tissues and tumors. When the Merlin was depleted in the EC, the researchers found that an excessive amount of non-functional vessels are formed in the tumor model, which enhances tumor necrosis and reduces tumor growth. Merlin-deficient mice implanted with human lung cancer showed an impressive 80.6% reduction in tumor growth after 3 weeks in comparison to regular mice. Likewise, adult vessels in the thyroid gland and small intestine showed an increased number of filopodia in Merlin-deleted mice in comparison to wild-type mice. Director KOH Gou Young of the Center for Vascular Research summarized, “Our study showed that Merlin is mainly localized at the cell membrane in vascular ECs, interacts with VEGFR2 and VE-cadherin, and plays a negative role in VEGFR2 intracellular downstream signaling by suppressing VEGFR2 internalization. As a consequence, endothelial Merlin can be seen as a gatekeeping regulator for tip EC induction in retinal sprouting angiogenesis during postnatal development, for formations of filopodia and sprouts in matured and established capillary ECs of adults, and for tip cell formation and proper tumor vessel construction in tumor vessels.”
Schematic diagram depicting roles of Merlin in different densities of VE-cadherin in each EC. Active Merlin forms a complex with VEGFR2 and VE-cadherin at relatively high VE-cadherin density, while inactive Merlin (S518) allows high VEGFR2 internalization in ECs with low VE-cadherin density, which leads to tip EC induction.
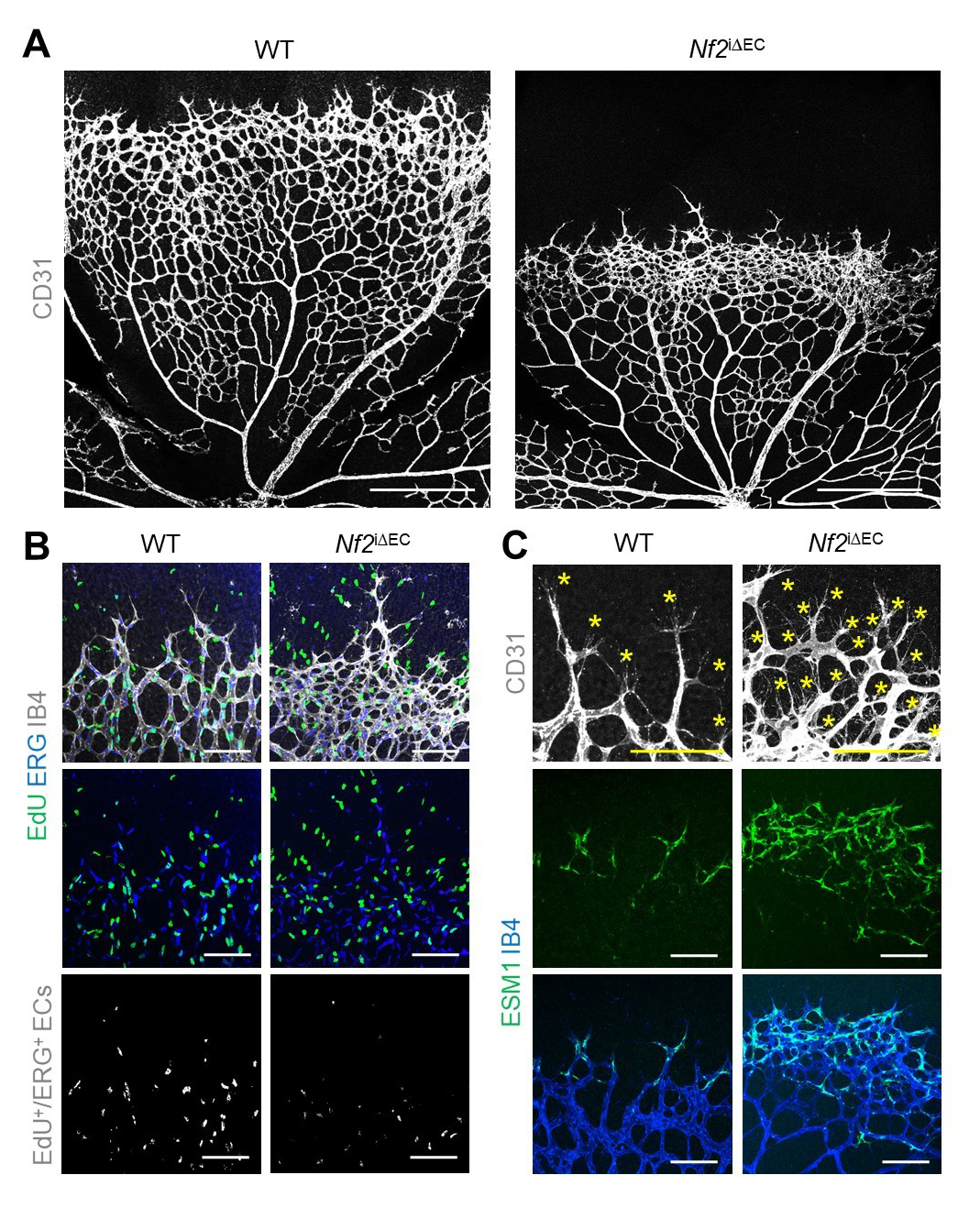
When Merlin was depleted, tip cells and filopodia at the vascular front were enhanced, but cell proliferation was reduced compared with WT mice (Fig.2 A-B). Endothelial cell-specific molecule-1 (ESM-1), which is a tip cell marker, was increased in Merlin-depleted mice compared with WT mice (Fig.2 C).
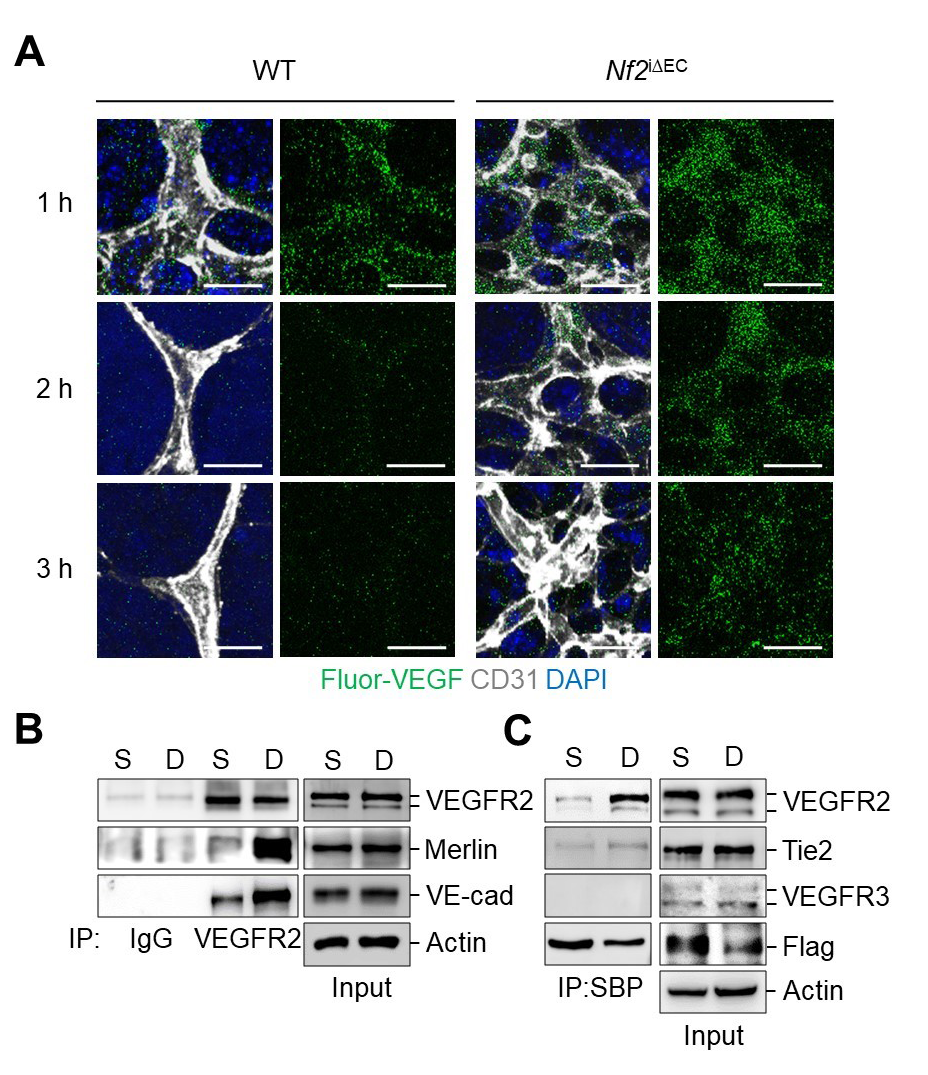
After Merlin inhibition in ECs, VEGF internalization at the vascular front was promoted compared with WT mice (Fig.3 A). Under dense cultured conditions (D), Merlin/VEGFR2/VE-cadherin interaction was enhanced while the complex was not associated under sparsely cultured conditions (Fig.3 B). Moreover, Merlin exclusively binds to VEGFR2 among other angiogenic receptor tyrosine kinase (Tie2, VEGFR3) (Fig.3 C). Notes for editors
- Reference
- Media Contact
- About the Institute for Basic Science (IBS)
|
|||
|
|
| Next | |
|---|---|
| before |
- Content Manager
- Public Relations Team : Yim Ji Yeob 042-878-8173
- Last Update 2023-11-28 14:20

55, Expo-ro, Yuseong-gu, Daejeon, Korea, 34126
Tel. +82-42-878-8114 | Fax. +82-42-878-8079 | E-mail. webmaster@ibs.re.kr
Copyright(c) IBS. All Rights Reserved.
